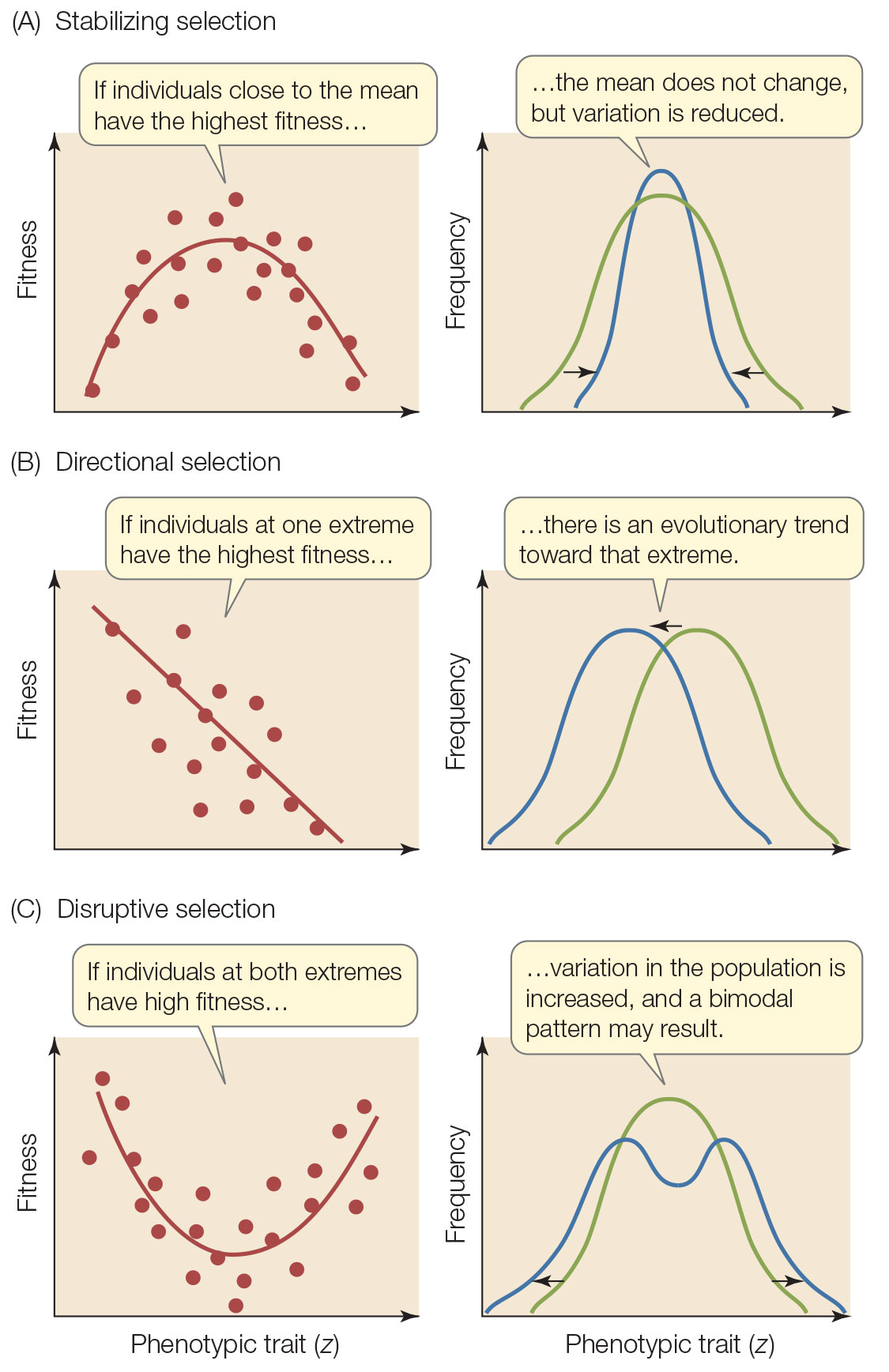
Вот эти работы Аксельрода и Гамильтона, я думаю, более интересны с точки зрения истории науки. Всё же парадигма группового отбора была тогда слишком сильной в эволюционной науке, а некоторым даже этого казалось мало:
Ю.Одум в монографии "Экология" писал(а):
В числе других Саундерс и Хо (Saunders, Но, 1976) утверждают, что сложность окружающего нас мира невозможно объяснить отбором только на видовом и организменном уровнях, поэтому главную роль в эволюции должен играть отбор на высших уровнях.
В этой книге автор немного касается также работ Аксельрода и Гамильтона. Взвешено, без критики, но честно намекает, что всё это вилами по воде писано :)
В закрытой теме я старался объяснить, что групповой отбор и родственный отбор (кин-отбор) --- это две кардинально разные вещи. Вилами по воде писан именно групповой отбор, это абстрактная гипотеза без известных (наблюдаемых или даже гипотетических) механизмов реализации этого самого "отбора на высших уровнях".
Работы Гамильтона и статья, написанная с Аксельродом, --- это про кин-отбор, то есть про отбор на уровне генов, который в большинстве случаев и с минимальными неточностями можно рассматривать как отбор на уровне организмов, индивидуальный отбор. Это значит, что теория классического дарвиновского индивидуального отбора --- это приближение к более точной теории отбора на уровне генов.
А групповой отбор --- это антидарвинизм, антинаука и плохая философия. Это то, о чем Докинз пишет в цитатах, которые Вы оставили в закрытой теме:
Р.Докинз в "Расширенном фенотипе" писал(а):
... хотя верно то, что желание обращать внимание на меньшие чем особь единицы, действующие во имя собственной выгоды возникает редко, но то же самое не всегда верно для единиц, бОльших особи. Годы, прошедшие со времён Дарвина, видели удивительное отступление от его сфокусированной на особи позиции -- впадение в сентиментально-бессознательный групповой отбор...
И только в последние годы, примерно совпавшие с запоздалым вхождением в моду собственных идей Гамильтона (Докинз 1979b), это сумасшедшее бегство было остановлено и перенаправлено. С тяжёлыми боями мы пробивались назад, изнурительно отстреливаясь от иезуитски искушённого и освящённого арьергарда новых групповых селекционистов, пока мы наконец не восстановили Дарвиновские позиции, которые я обозначаю меткой "эгоистичный организм"; позиции, которые в их современной форме доминируют в концепции "итоговой приспособленности".
Р.Докинз в "Расширенном фенотипе" писал(а):
Эгоистичный организм, и эгоистичный ген с его расширенным фенотипом -- есть два представления одного и того же куба Неккера.
Групповой отбор
не является еще одним представлением "одного и того же куба Неккера", а индивидуальный отбор (эгоистичный организм) и кин-отбор (эгоистичный ген) являются. Групповой отбор --- это, заостряя, просто сентиментально-бессознательная ерунда. Бессознательная, потому что никто из "групповиков" не нашел механизм этого самого "отбора на высших уровнях", на уровнях выше индивидуального.
Устарел именно групповой отбор. Всё, что он дал, --- это четкое понимание, что теория индивидуального отбора работает не всегда (истинный альтруизм). Но это вопросы без ответов. Ответы дала теория отбора на уровне генов.
Одум, насколько я вижу по 8-ой главе его книги, считает, что групповой отбор живее всех живых. И к тому же, он, кажется, не видит важнейшей разницы между групповым и родственным отбором --- обычное дело даже для биологов. (Все это усугубляется терминологической путаницей в биологической литературе.)
Что касается Вашей последней ссылки на Википедию, обратите внимание на "Категории", к которым она относится. Это не статья о самой "эволюции кооперации", она написана по поводу книги / статей того периода. (А обилие какой-то философии с идеологией не нравится в той статье не только мне, похоже :)
Если смущает статья в Вики, то могу заверить, что в статье Гамильтона и Аксельрода идеологии нет. Только биология и простая математика. Вместе со статьёй Триверса (Trivers, "The Evolution of Reciprocal Altruism" (1971)) --- это ясное объяснение того, как и при каких условиях может возникнуть межвидовая кооперация: нет почти никаких отличий между формированием взаимного "альтруизма" (а точнее, взаимной кооперации) внутри вида и между видами. Есть совпадение предсказаний с наблюдениями.
По поводу моих слов об "энергоэффективности". После беглого прочтения упомянутого выше двухтомника по экологии я решил от них отказаться. Не то чтоб это было совсем ошибочно, но слишком много нужно сделать оговорок, чтоб так говорить.
Все-таки я думаю, что "энергоэффективность" --- это ошибочный подход. Энергетические факторы являются общими ограничителями, но не являются формирующими причинами той или иной эволюционной стратегии.
(Оффтоп)
Используя метафору глины, можно сказать, что свойства глины ограничивают возможности ее применения, но не определяют, чем станет эта глина внутри границ возможного: тарелкой, кувшином или статуэткой. Ясно, что из глины не вылепишь радиоприемник или самолет. (Вылепить можно только визуальный образ приемника или самолета.) Так же и энергетические ограничения --- это широкие границы, внутри которых формируются разные эволюционные стратегии.
Но это все философия: различение материальной и формальной (формирующей) причины у Аристотеля, например.
Теперь конкретика. Вы написали:
Рассмотрим умозрительно 2 вида животных, конкурирующих в одной общей экосистеме. Если (при прочих равных!) один из этих видов способен получить необходимую для выживания энергию с меньшими затратами, то он получит преимущество в этой конкурентной борьбе.
Давайте определим "прочие равные" и "прочие неравные".
Прочие равные: особи обоих видов плодятся в одно и то же время (ровно в 00 часов 00 минут каждых суток), производя по одному отпрыску. Отпрыск немедленно умирает, если для него не находится энергетических ресурсов (пищи). Если пища есть, отпрыск достигает зрелости за сутки и производит своего потомка в 00 часов и 00 минут. Особи живут 10 суток. Умирая они передают свою энергетическую нишу своим непосредственным отпрыскам.
Прочие неравные: особь первого вида потребляет 1 единицу пищи в сутки, особь второго --- 2.
Среда производит 192 единицы пищи в сутки. В первые сутки, в 00 часов 00 минут, мы пересаживаем в среду одного новорожденного отпрыска первого вида и одного новорожденного отпрыска второго вида. Через 6 суток и далее у нас будут жить 64 особи первого и 64 особи второго вида, полностью утилизирую весь доступный поток энергии. Никакой конкуренции здесь нет.
Чтобы допустить какую-то конкуренцию, нужно обогатить модель. Надо добавить, например, борьбу за территорию и связь между контролируемой территорией и доступом к энергии. Или добавить борьбу через скорость или периодичность размножения. Или допустить поедание особями друг друга (или любую другую перекачку энергии из одних особей в другие). Обязательно нужно добавить связи между всем этим и числом потомков. И так далее. Всё это может происходить миллионами разных путей, которые уже реализованы в природе, или которые эволюция отыщет в будущем. Сомнительно, чтобы в такой сложной модели можно было бы установить какие-то корреляции между как угодно измеренной энергоэффективностью особей, их совокупностей и устойчивыми эволюционными стратегиями. Сама природа является "реализацией" такой модели. А она очень разнообразна по энергоэффективности особей, разнообразны стратегии. И, главное, разнообразие в масштабах миллиардов лет только нарастало.
В теоретической биологии пошли другим путём, спустились сразу к истоку: Гамильтон ввел понятие "итоговая (инклюзивная) приспособленность" (inclusive fitness). Это число потомков, имеющих тот же генотип. Здесь не уточняется какие конкретно действия приводят к увеличению или уменьшению этой приспособленности. Их миллионы. Важно только, что взаимодействующие организмы могут как-то повлиять на приспособленность друг друга. А дальше в ход идут инструменты теории игр.
Эти мои размышления подводят к такому выводу: Каждому конкретному взаимодействию организмов можно приписать какую-то энергетическую меру. (И, конечно, не одну.) Но, скорее всего, для всей совокупности живых существ никакой корреляции между энергоэффективностью и изменением приспособленности не будет. Например, кооперация может быть и энергоэффективной, и энергозатратной для кооператоров, но она всегда предпочтительна с точки зрения инклюзивной приспособленности, если является устойчивой эволюционной стратегией.





{kind=link}