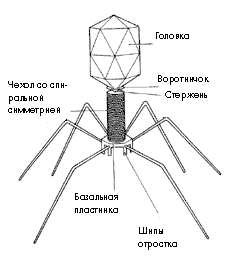
Возможный механизм возникновения конъюгации (КОНЪЮГАЦИЯ - способ переноса генетического материала
от одной бактериальной клетки к другой).
Кратко изложу известный из литературы вариант возникновении жизни на Земле.
Рибоза — это сахар с пятью атомами углерода, который лежит в основе нуклеотидов, формирующих РНК и ДНК.
Реакция Бутлерова — автокаталитическая реакция синтеза из формальдегида CH2O различных сахаров, в том числе и рибозы, в слабощелочных водных растворах в присутствии ионов металлов.
Вблизи вулканов, где синтезируется формальдегид, в скальных расщелинах могли образовываться колонии молекул сахаров-автокатализаторов - прото-РНК на основе рибозы.
Подробнее о теории возникновения жизни можно прочитать в статье акад. В. Н. Пармона по адресу:
http://elementy.ru/lib/25618/25620 Первичная наследственность была в виде однонитчатой молекулы т.е. в начале были прото-РНК. Колонии прото-РНК “приручили” белки и научились строить оболочку. Так возникли одноклеточные - прото-бактерии.
Питание - через поглощение себе подобных.
Размножение, как результат невозможности удержать под одной оболочкой всю накопившуюся массу.
Пополнение информации происходит через включение в генетический аппарат поглощенных из окружающей среды свободных цепочек прото-РНК, оставшихся после гибели таких же одноклеточных..
Возникают прото-бактериофаги, паразитирующих на прото-бактериях.
Современные фаги
Видео “Бактериофаг инфицирует бактерию” смотри по адресу:
http://www.youtube.com/watch?v=41aqxcxsX2w&feature=player_embedded Нуклеиновая кислота бактериофага, введённая в бактериальную клетку, управляет клеточными механизмами и программирует синтез новых фаговых частиц в количестве 100…1000 единиц. Затем наступает лизис - растворение оболочки бактерии, и фаговые частицы готовы к следующему циклу заражения. В случае т.н. умеренных бактериофагов лизис не наступает. Умеренные фаги интегрируются в геном клетки-хозяина или существуют в виде плазмид.
Рассмотрим гипотетический случай: некоторые из тем не менее образовавшихся умеренных прото-бактериофагов в первой бактерии-хозяине могут проколоть изнутри оболочку не только бактерии-хозяина, но и дотянуться до оболочки второй бактерии находящейся рядом, и затем инфицировать ее. Такой гипотетический механизм инфицирования второй бактерии бактериофагом, который находится в первой бактерии, может закрепиться. Это возможно, например, если геном бактериофага был предварительно интегрирован с геномом первой бактерии, а затем, обогащенный частью генома бактерии, встроен в фаговую оболочку. Т.е теперь геном бактериофага состоит из части генома бактерии и генома первичного бактериофага. Вторая бактерия-реципиент в результате инфицирования может получить перспективы, как для выживания, так и для повторения инфицирования следующей, третьей бактерии, которая так же может продолжит начатую цепочку передачи генетической информации. Этот механизм, возможно, мог дать начало существующей ныне бактериальной к о н ъ ю г а ц и и, а в конце архея, ~2,5 млрд. лет назад, разделению организмов на два пола.
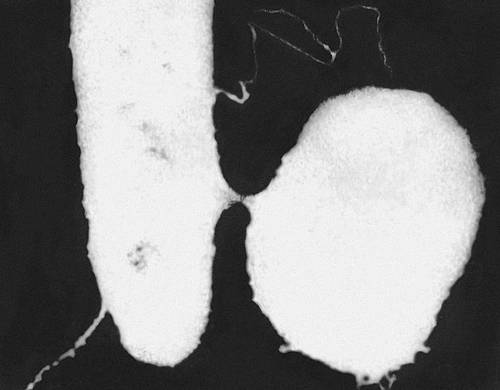
Конъюгация бактерий: между палочковидной клеткой-донором и округлой клеткой-реципиентом виден тонкий соединительный канал по которому происходит перенос генетического материала.
Т.е., возможно, у истоков разделения живых организмов на два пола были вирусы.